2025年5月6日,国际综合期刊Journal of Advanced Research(中科院1区TOP,IF=11.4)在线发表了题为Heterologous expression of the barley-specific HvbZIP87 transcription factor in wheat enhances broad-spectrum disease resistance with balanced yield的研究论文。文章报道了利用大麦特异转录因子HvbZIP87提升小麦广谱抗病水平并维持产量平衡的研究,为小麦抗病遗传改良提供了创新性基因资源和技术支撑。河北农业大学王逍冬教授和西北农林科技大学王晓杰教授为论文通讯作者,河北农业大学已毕业博士生赵淑清和西北农林科技大学王宁教授为论文第一作者。中国工程院院士、西北农林科技大学康振生教授对研究工作给予了指导。研究工作得到了国家重点研发计划(2023YFD1201000/2021YFD1401000)、河北省杰出青年基金(C2022204010)、中央引导地方项目(236Z6501G)、石家庄驻冀高校重点研发计划(241490012A)、华北作物改良与调控国家重点实验室自主课题(NCCIR2024ZZ-5/23567601H)和河北省旱碱麦产业技术体系(HBCT2024030206)的资助,并已授权国家发明专利1项(ZL202011299785.9)。
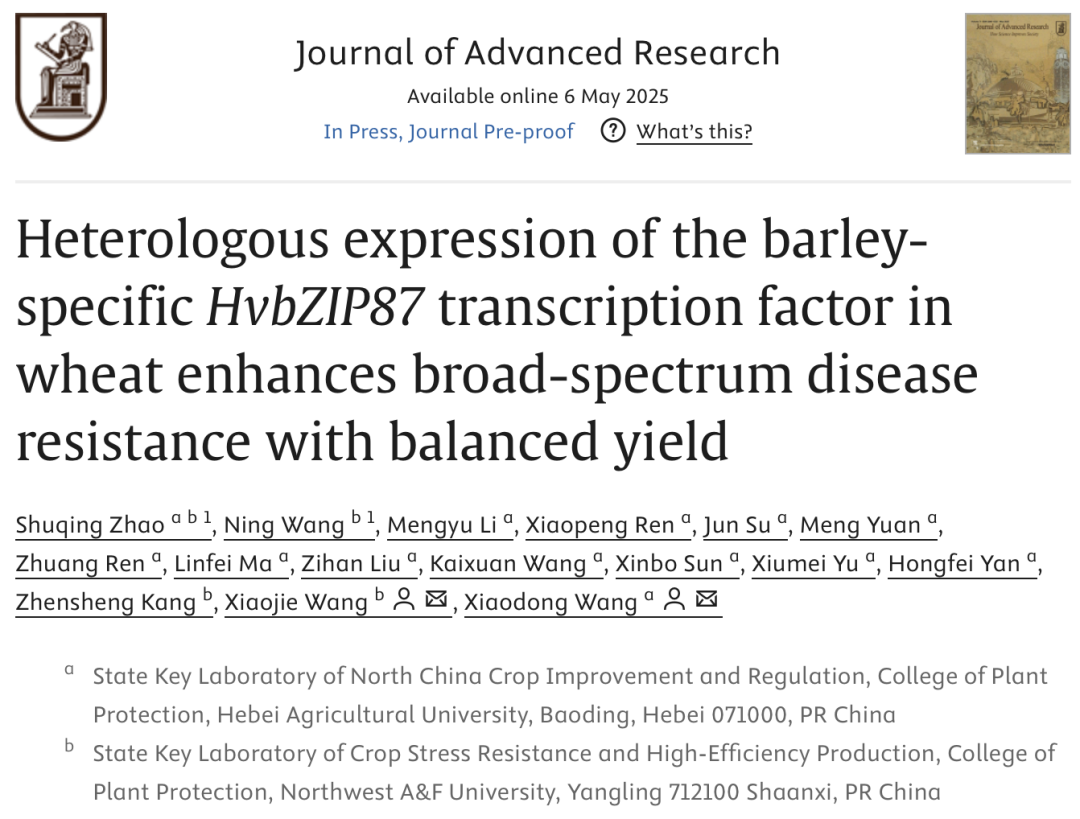
小麦作为我国的重要粮食作物,其稳定增产直接关系到我国的社会经济发展。然而,受到单一小麦品种大面积种植、全球气候变暖等因素影响,小麦条锈病、叶锈病、白粉病等叶部真菌病害在我国不同麦区均有发生与流行,严重威胁我国小麦生产与粮食安全。一般认为,植物激素水杨酸(SA)在作物抗锈菌、白粉菌等活体营养型病原真菌过程中起关键作用。而同样由SA介导的系统获得抗性(SAR),是植物自身应对初生病原菌侵染诱导产生的抗性反应,具有针对多种次生病原菌的广谱抗性特征。如何利用植物SAR过程关键基因提升作物抗病水平,是植物免疫学领域从抗病分子机制研究到关键基因开发利用的重要科学问题。本研究通过丁香假单胞菌DC3000诱导大麦系统获得抗性(SAR)反应,全基因组分析发现bZIP转录因子基因Novel07221被显著诱导表达,且在HvNPR1沉默株系中表达量进一步升高。序列比对及进化分析显示,该蛋白与大麦HvbZIP87(MLOC_12585)和HvbZIP10(ASU50458)高度同源,故暂命名为HvbZIP87。系统发育分析表明其与小麦TaNPR1互作的TGA转录因子不同源,且小麦基因组中未发现直系同源基因,提示该基因可能为大麦特有。HvbZIP87基因ORF全长663 bp,编码221个氨基酸(预测分子量24.3 kDa),含核定位信号(NLS,71-77aa)和bZIP结构域(67-132aa)。亚细胞定位实验证实:通过农杆菌介导的烟草瞬时表达体系,GFP-HvbZIP87及HvbZIP87-GFP融合蛋白均定位于细胞核并与DAPI核染色共定位;NLS突变体(71-77aa替换为7A)导致核定位缺失,证实该区段为核定位关键位点。研究揭示HvbZIP87为大麦特异性bZIP转录因子,其核定位功能依赖特定NLS序列。
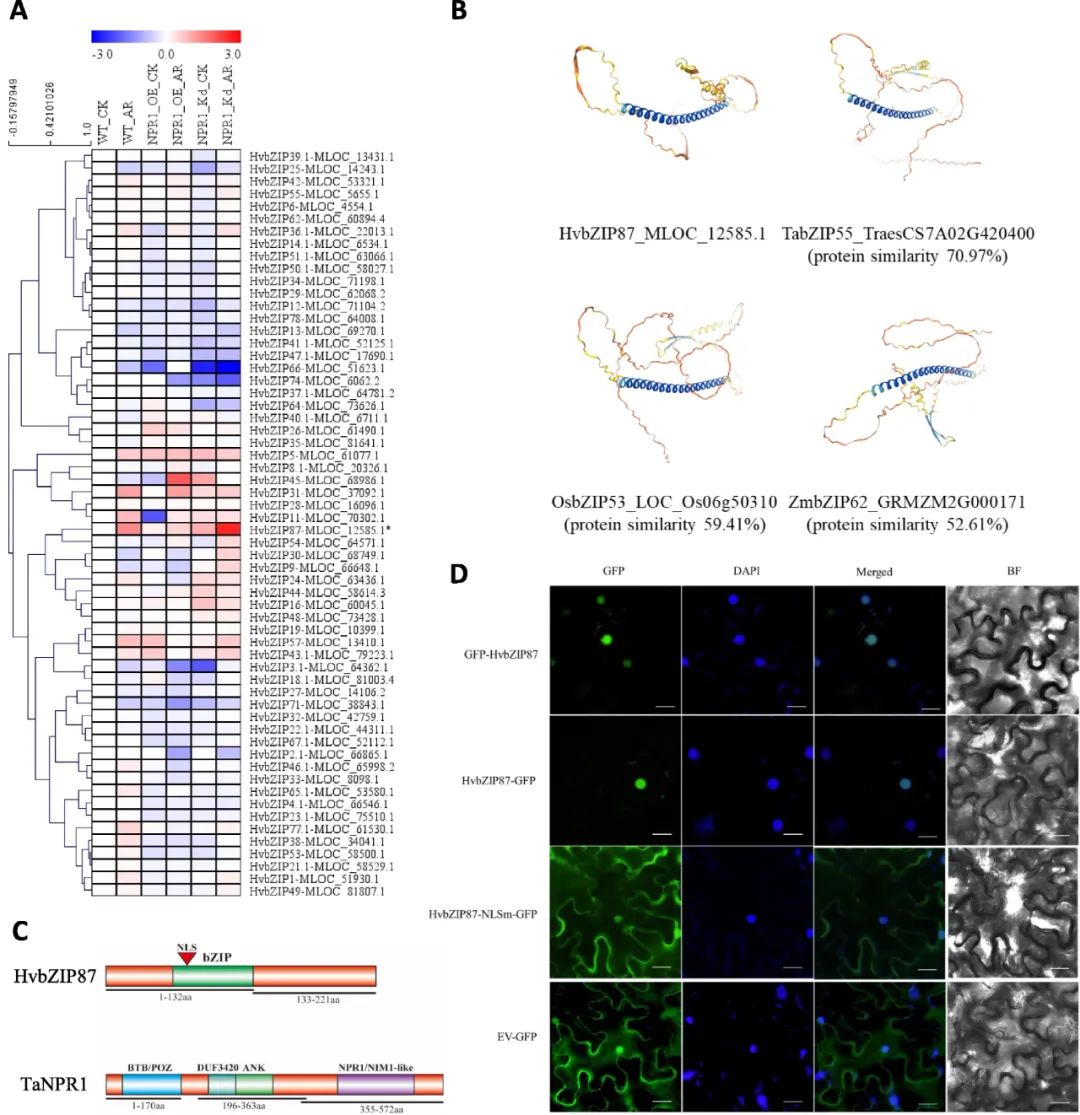
为探究HvbZIP87功能,构建了HvbZIP87-pLGY-02小麦转基因载体,在济麦JW1中过表达该基因(HvbZIP87-OE),并通过qRT-PCR验证其表达量为野生型(WT)的8-12倍。在叶尖注射丁香假单胞菌DC3000诱导SAR反应后,相邻区域接种稻瘟病菌。结果显示,HvbZIP87-OE小麦病斑面积显著小于WT植株,表明过表达HvbZIP87可增强SAR反应及对次生病原菌的抗性。通过qRT-PCR检测病程相关蛋白(PR)基因(TaPR1a、TaPR2、TaPR4b)表达,发现DC3000诱导后,HvbZIP87-OE小麦中PR基因表达水平显著高于WT,证实HvbZIP87过表达激活了小麦的SAR反应及防御相关转录应答。
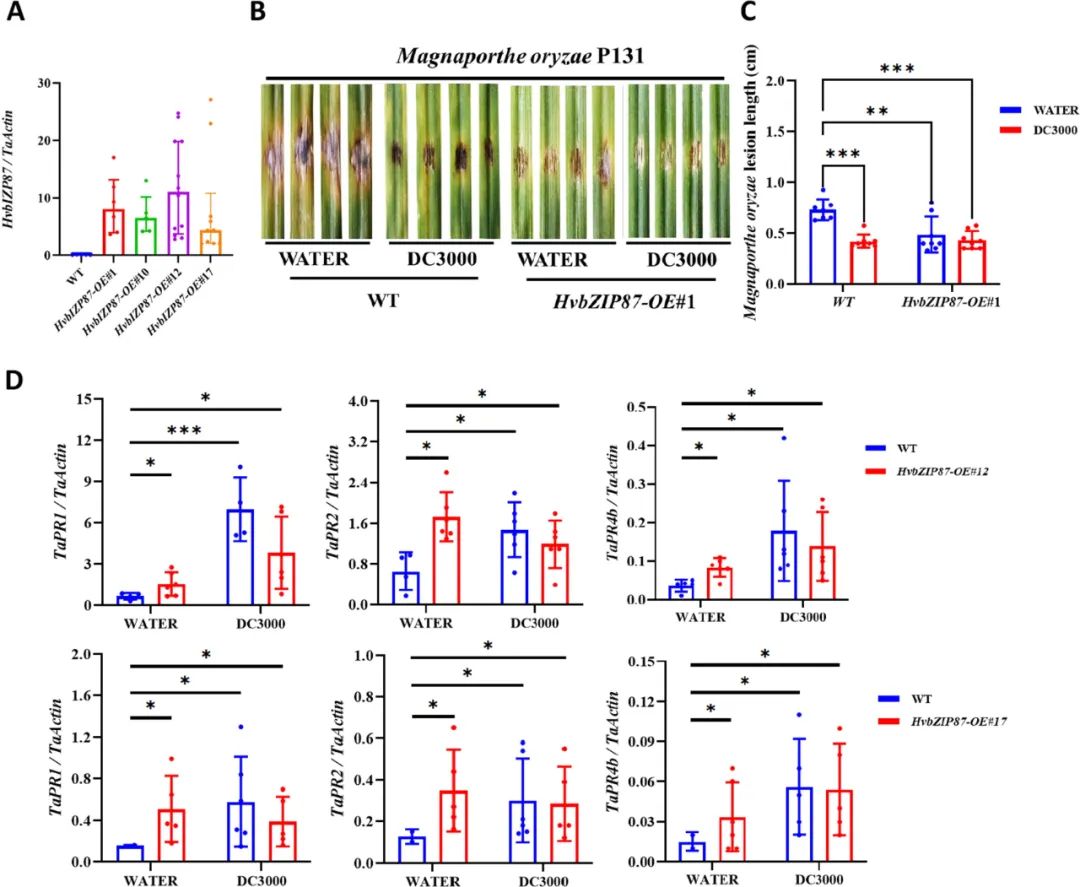
为验证HvbZIP87过表达对小麦抗病性的影响,对HvbZIP87-OE转基因植株及野生型(WT)接种多种病原菌并检测抗病表型。结果表明:接种条锈菌(CYR32)和叶锈菌(THTT)后,转基因植株锈孢子堆面积占比显著低于WT,且叶锈菌感染症状减轻;接种根腐叶斑病菌后,转基因植株病斑面积明显小于WT;茎基腐病菌感染则显示转基因植株病斑颜色更浅、长度更短。
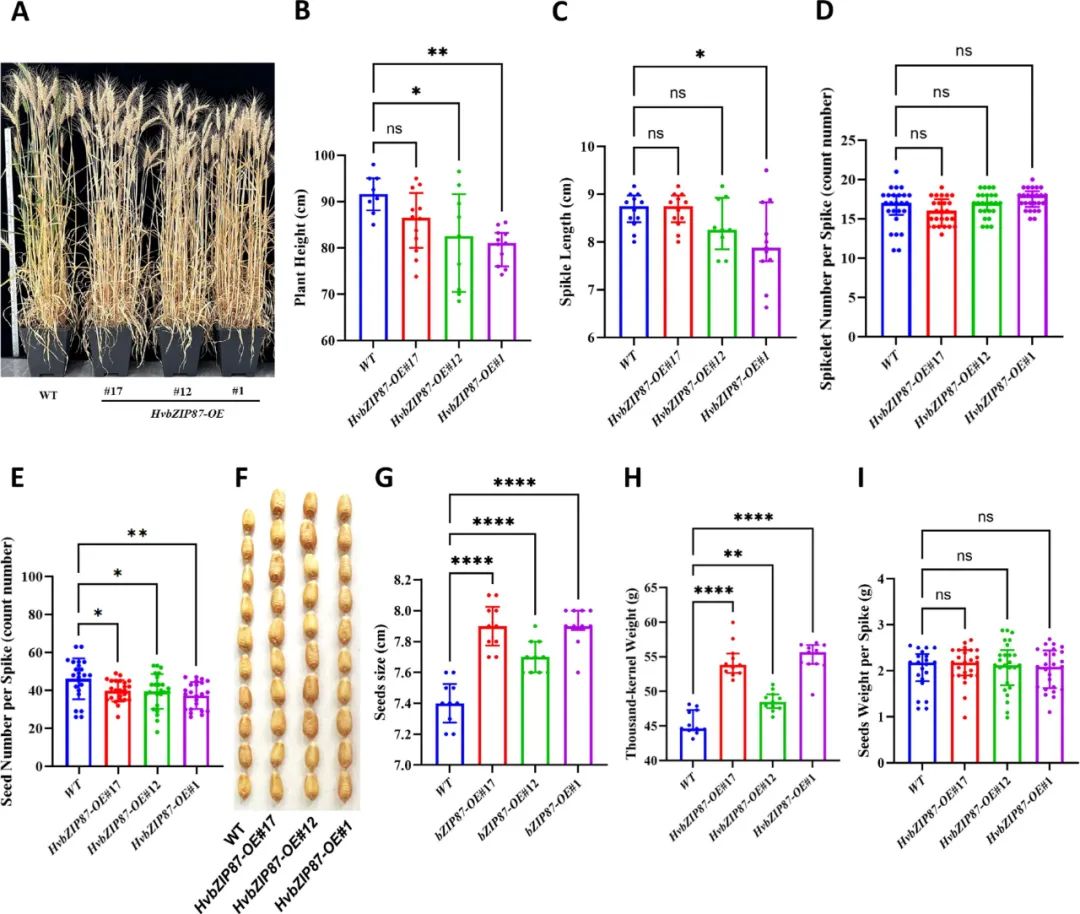
为了验证在小麦中过表达HvbZIP87是否会影响农艺性状,我们在温室条件下对HvbZIP87-OE小麦转基因和WT植株的株高、穗长、小穗数、穗粒数、种子大小和千粒重等农艺性状进行了统计。与WT植株相比,HvbZIP87-OE小麦转基因株高略低、穗长略短,小穗数与WT植株几乎一样,HvbZIP87-OE小麦转基因穗粒数略少于WT植株,但是HvbZIP87-OE小麦转基因收获种子的千粒重与种子饱满程度都高于WT植株,说明在小麦中过表达HvbZIP87不会影响小麦产量。
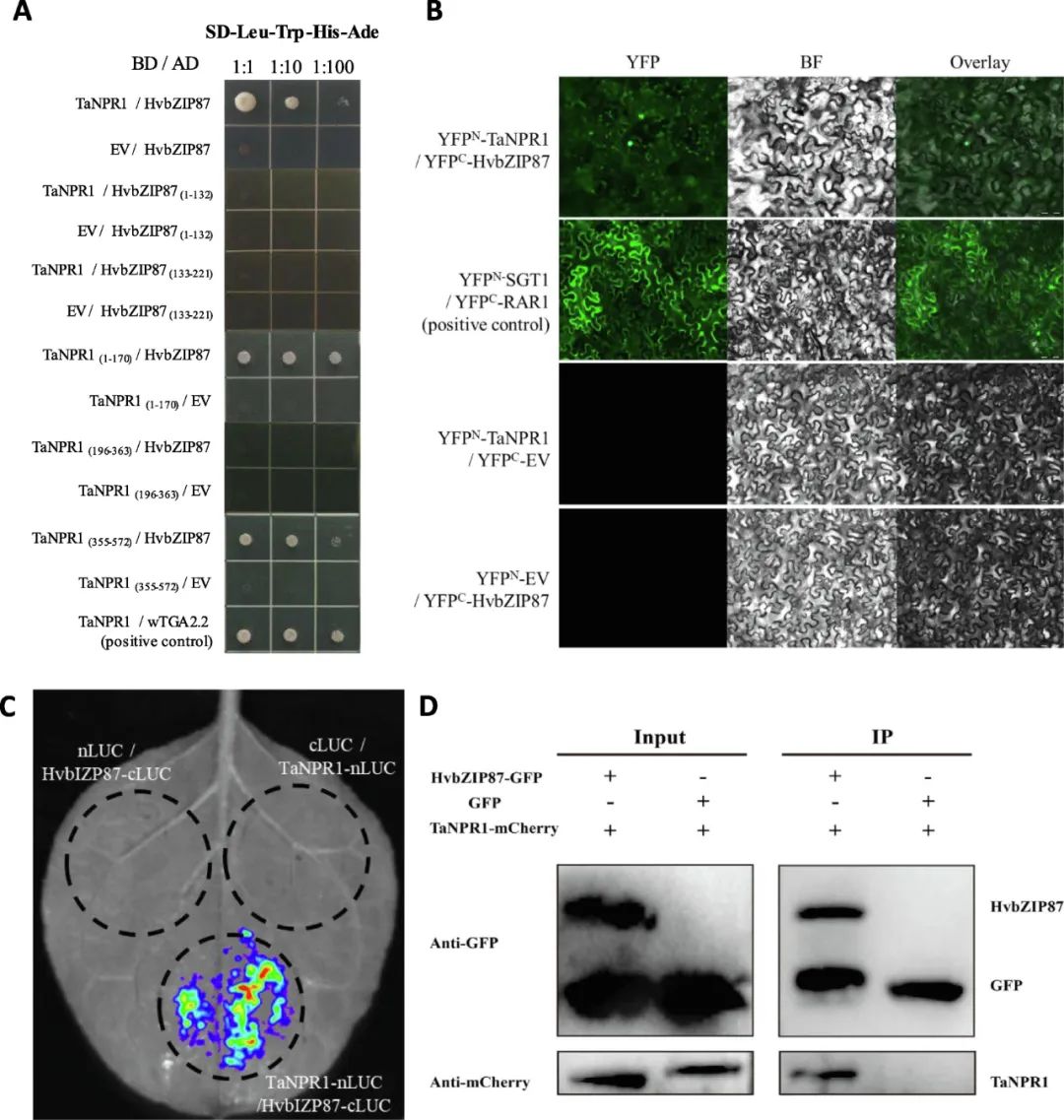
为探究HvbZIP87与TaNPR1的蛋白互作关系,通过多维度实验验证了两者相互作用及其功能意义:首先,蛋白共定位实验显示TaNPR1-GFP与HvbZIP87-mCherry在细胞核中共定位,提示潜在互作;进一步采用酵母双杂交技术,证实HvbZIP87与TaNPR1的BTB/POZ(1-170aa)及NPR1/NIM1-LIKE(355-572aa)结构域直接互作,且该互作不依赖HvbZIP87的bZIP保守结构域(67-132aa)。随后,双分子荧光互补(BiFC)及荧光素酶互补实验(LCA)在植物体内外均检测到核内荧光信号,明确互作发生在细胞核;最终,免疫共沉淀(Co-IP)实验通过农杆菌瞬时表达体系,进一步在植物体内验证了HvbZIP87-GFP与TaNPR1-mCherry的特异性互作。值得注意的是,与经典NPR1-TGA互作模式不同,HvbZIP87可能通过非ANK结构域介导的全新机制与TaNPR1相互作用,为揭示NPR1调控网络提供了新视角。
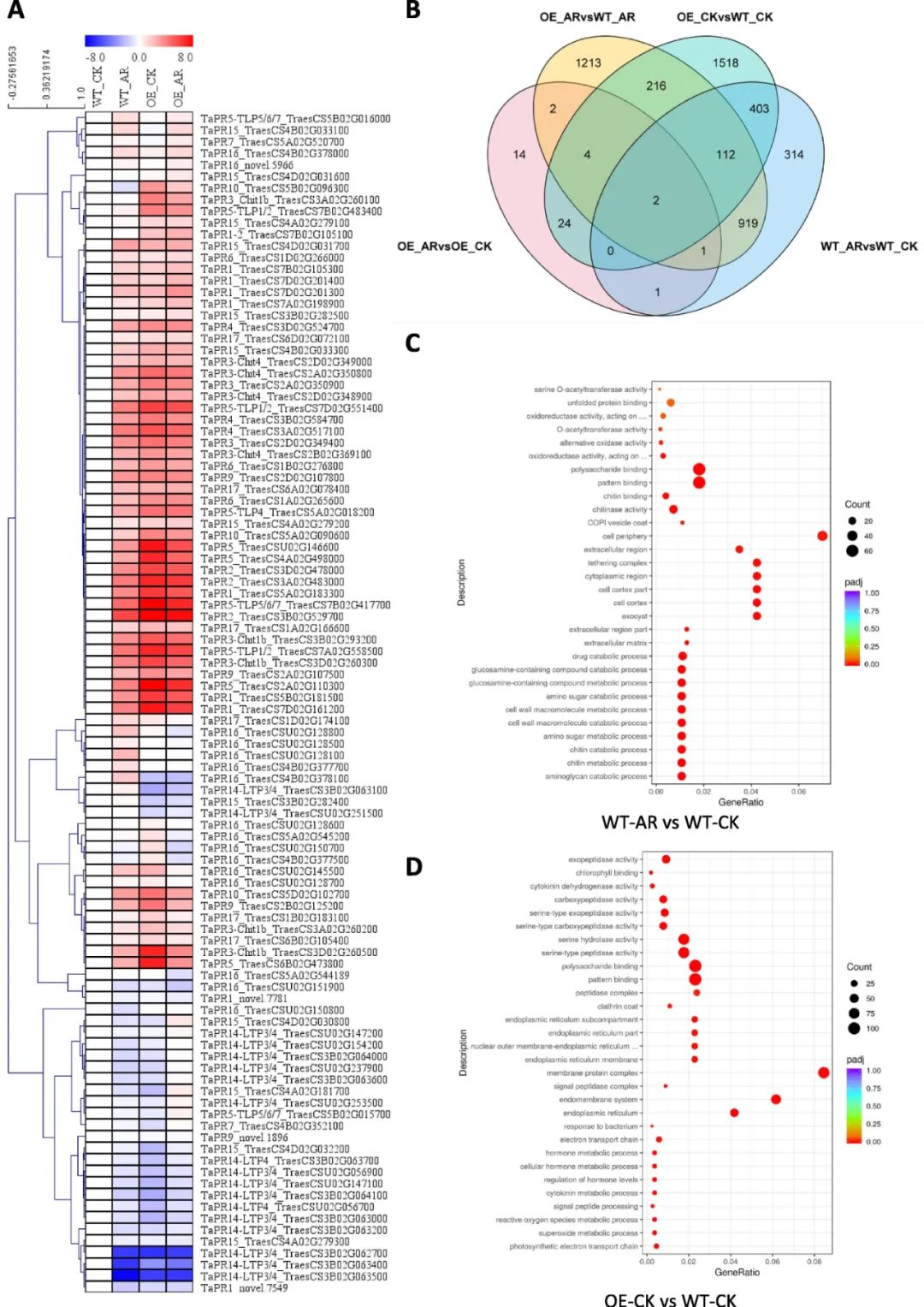
为解析HvbZIP87抗病机制,对转基因及野生型小麦进行转录组测序分析,通过丁香假单胞菌DC3000诱导后,提取叶片RNA进行12 Gb Illumina测序,共鉴定116,815个转录本,生物学重复相关性高(R²>0.92),数据可靠。结果显示:约2/3病程相关蛋白(PR)基因在野生型中受病原菌诱导上调,而在转基因植株中,无论清水或病原菌处理均检测到PR基因上调,且PR14基因表达均被抑制,表明HvbZIP87过表达可独立于病原菌部分激活系统获得抗性(SAR)相关转录响应。以“|Log2FC|>1且q为阈值,发现野生型病原菌处理组(WT_AR)与对照组(WT_CK)存在显著转录差异,且与转基因组清水处理(OE_CK)存在部分重叠DEGs。基因本体(GO)富集分析显示“多糖结合”和“模式识别”相关功能显著富集。KEGG通路分析表明,野生型中病原菌激活钙信号、WRKY转录因子及效应蛋白触发的免疫(ETI),而转基因植株中部分钙信号及ETI通路被特异性激活,提示HvbZIP87可能通过调控钙信号及ETI通路增强抗病性。
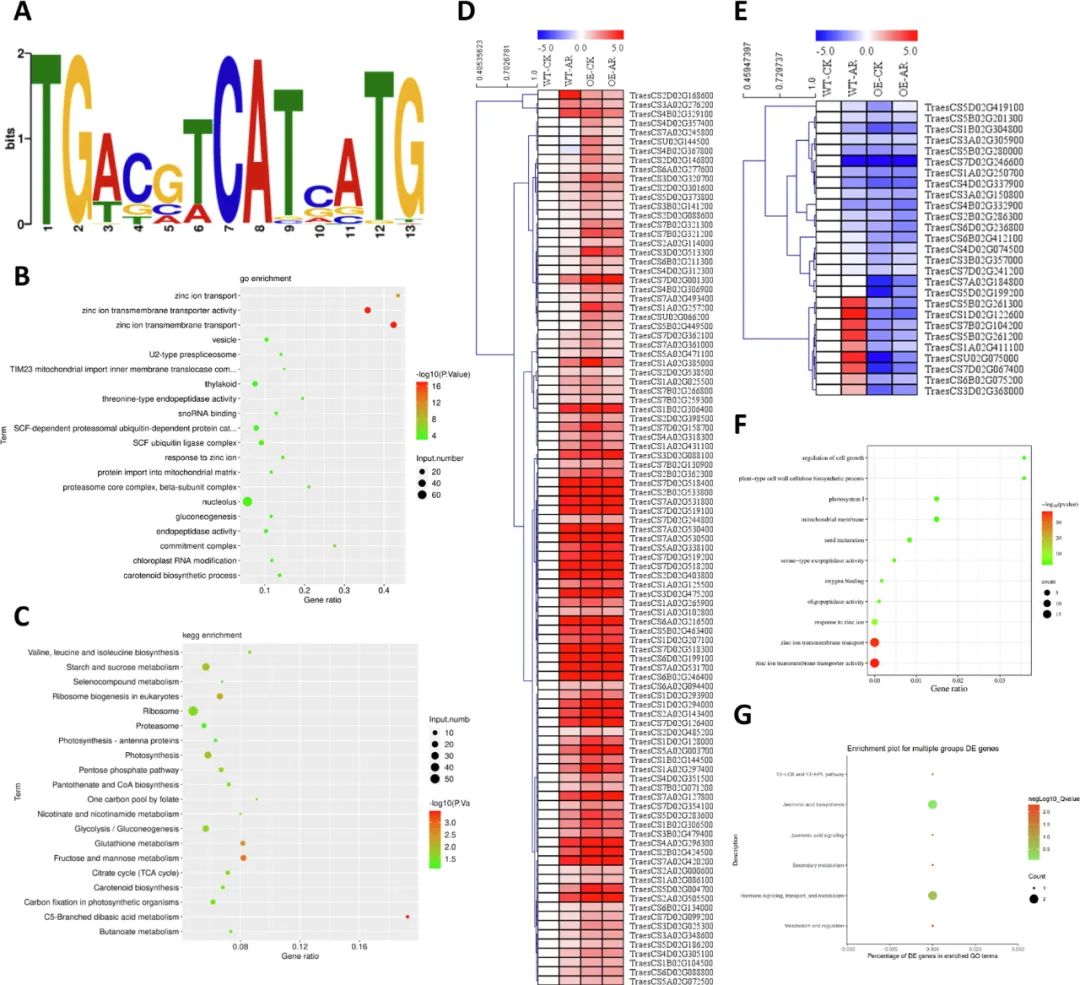
通过KEGG通路分析揭示HvbZIP87对植物激素信号通路的调控作用:在野生型中,丁香假单胞菌DC3000诱导SAR反应后,吲哚乙酸(IAA)、细胞分裂素(CTK)、脱落酸(ABA)、乙烯(ETH)、水杨酸(SA)等通路均出现差异表达;而转基因植株中,赤霉素(GA)和茉莉酸(JA)通路基因表达模式发生显著改变。激素定量分析显示,过表达HvbZIP87抑制了SA和GA的积累,但显著增强ABA含量。进一步通过DAP-seq技术鉴定出HvbZIP87直接结合的DNA元件(“TGACGTCATCATG”),其靶基因富集于“锌离子跨膜转运”和“C5-支链二元酸代谢”通路。转录组分析表明,193个结合基因在转基因植株中上调表达,且这些基因同样显著参与“锌离子转运”过程,提示HvbZIP87可能通过调控锌离子平衡及激素信号网络增强植物抗病性。
通过酵母双杂交筛选发现HvbZIP87与72个候选蛋白存在互作,其中重点鉴定了MYC类转录因子TaMYC2。验证表明,HvbZIP87与TaMYC2在细胞核内直接互作,且TaMYC2在转基因小麦中表达受抑制。进一步研究发现,TaMYC2定位于细胞核,并通过病毒介导的基因沉默(VIGS)技术证实其沉默显著降低小麦条锈菌侵染后的孢子堆面积,表明TaMYC2在植物防御反应中发挥负调控作用。

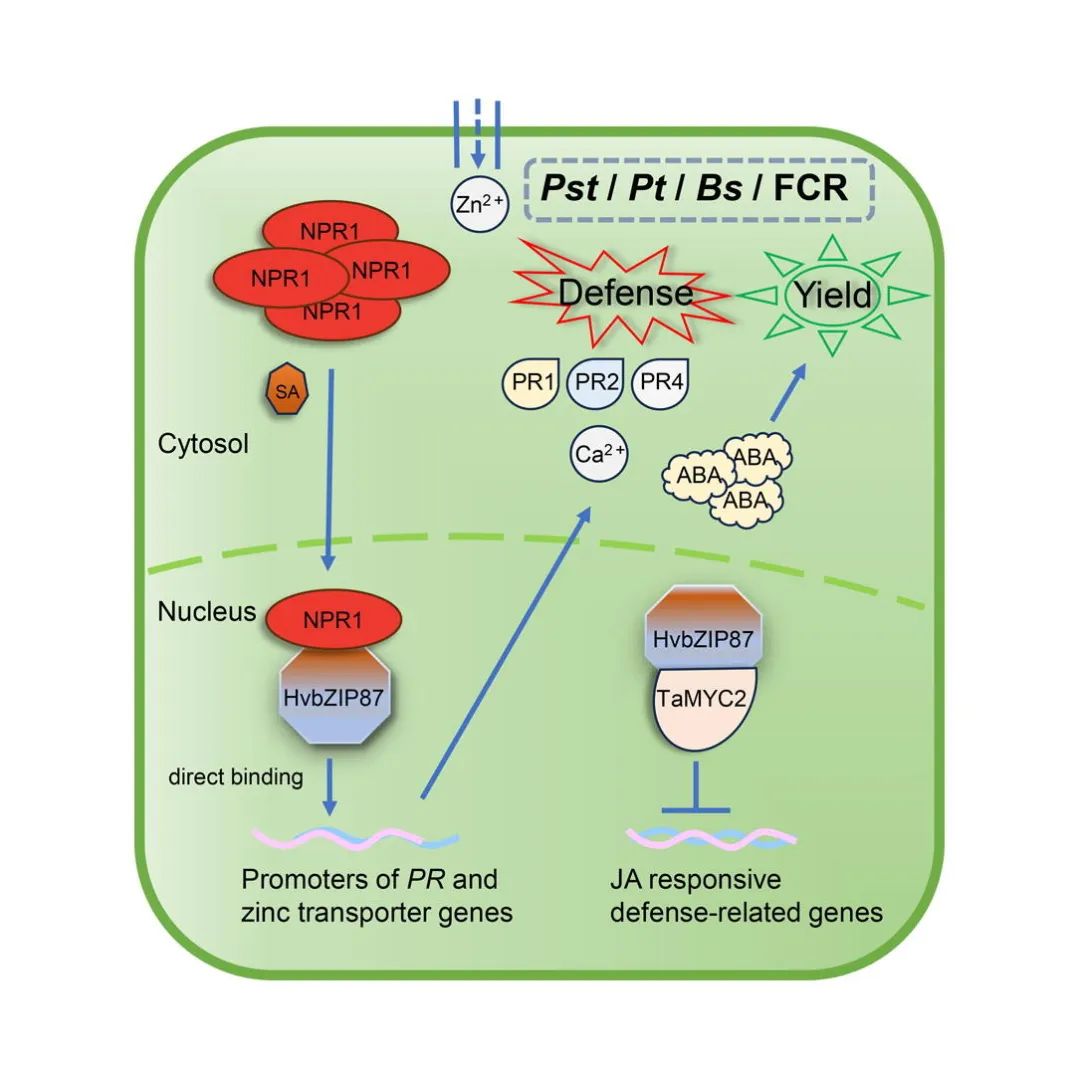
综上所述,过表达HvbZIP87基因的小麦通过多途径协同作用实现抗病与产量的平衡。HvbZIP87通过直接调控PR基因表达激活防御反应,同时与抗病核心调控因子TaNPR1核内互作,增强对多种病原菌的广谱抗性。转基因小麦在保持产量的前提下实现种子膨大,表明HvbZIP87的抗病功能未对农艺性状产生负面影响。HvbZIP87既通过锌转运相关基因调控离子平衡,又通过抑制防御负调控因子TaMYC2的功能,形成“激活-抑制”双效调控网络,揭示了其作为抗病枢纽的分子基础。
小麦族多组学网站:http://wheatomics.sdau.edu.cn
投稿、合作等邮箱:shengweima@icloud.com
(转自:小麦研究联盟)