由条形柄锈菌小麦专化型(Puccinia striiformis f. sp. tritici)引发的小麦条锈病是一种极具危害性的气传性真菌病害,通常会造成5%—30%的产量损失,严重时甚至绝收,对小麦安全生产构成重大威胁。种植抗病品种是防治该病害最经济有效的途径之一。条锈菌为活体营养专性寄生真菌,通过吸器向小麦细胞分泌效应蛋白,操纵植物免疫促进侵染。阻止病原菌对寄主的操纵,是创制抗病材料的切实可行途径。深入解析小麦与条锈菌互作机制,挖掘病菌致病关键基因和小麦免疫调控因子,可为小麦抗病遗传改良提供可利用基因与理论依据。
丝裂原活化蛋白激酶(MAPK)级联反应通过将植物细胞外的信号转导至细胞内,在植物先天免疫中发挥着关键作用,但在植物中仅有少数完整的MAPK级联反应被证实能够赋予对病原菌的抗性。鉴于MAPK级联反应在植物免疫中的重要作用,病原菌进化出了多种效应蛋白,靶向MAPK级联反应,从而操控宿主的免疫反应并促进自身的侵染。
近日,JIPB在线发表了西北农林科技大学作物抗逆与高效生产全国重点实验室/植物保护学院植物免疫团队题为“Wheat MAPK cascade mediates SGT1 nuclear entry targeted by a stripe rust effector”(全文链接https://doi.org/10.1111/jipb.13888)的研究。该研究鉴定出了一个有助于小麦抗条锈病的TaMKK2-TaMAPK6-TaSGT1信号级联反应,发现小麦条锈菌效应蛋白HASP215能够靶向TaMKK2互作,破坏TaMKK2-TaMAPK6-TaSGT1磷酸化级联反应,抑制寄主免疫,促进条锈菌侵染致病。
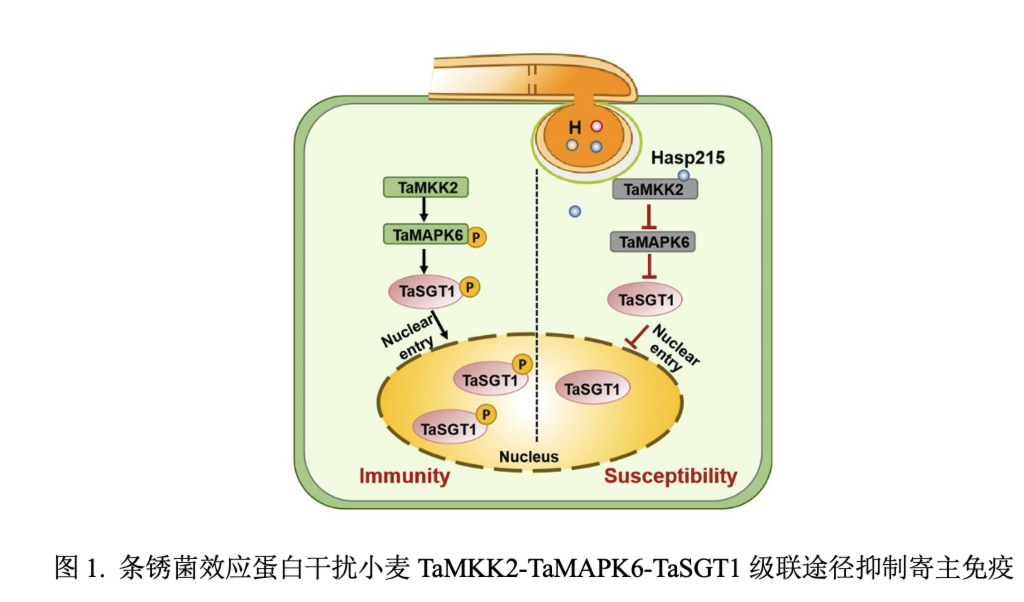
本研究鉴定到了一个小麦丝裂原活化蛋白激酶(MAPK)激酶(TaMKK2),利用基因敲除突变体和过表达转基因小麦,证明了TaMKK2是小麦抗条锈病的重要正调控因子。TaMKK2能够与TaMAPK6相互作用并使其磷酸化,而TaMAPK6能够磷酸化核心免疫调节因子TaSGT1(Skp1的G2等位基因抑制子),从而增强TaSGT1进入细胞核,赋予小麦对条锈病的抗性。为对抗小麦的防御,小麦条锈菌通过其特化侵染结构吸器向寄主细胞分泌毒性效应蛋白HASP215。HASP215靶向TaMKK2,通过干扰TaMKK2-TaMAPK6互作抑制TaMAPK6的磷酸化,致使TaMAPK6介导的TaSGT1磷酸化及细胞核转运受抑,最终导致小麦免疫被削弱。综上,该研究阐明了TaMKK2-TaMAPK6级联反应通过调节关键免疫成分TaSGT1正调控小麦抗病性,而HASP215作为锈菌关键效应蛋白直接抑制这一信号级联反应的小麦—条锈菌攻防机制。
攻击植物MAPK或SGT1已被病原体效应蛋白广泛采用。与之不同的是,该研究表明,一种锈菌效应蛋白靶向小麦免疫信号转导中的TaMKK2-TaMAPK6-TaSGT1模块,为深入理解MAPK级联反应在作物免疫中的作用,及专性活体营养型真菌的致病机制提供了新的见解,有助于更全面地理解植物免疫系统以及条锈菌的侵染途径。基于此,我们构建了稳定的Hasp215 RNA干扰和TaMKK2、TaSGT1过表达转基因植株,显著提高了小麦对条锈病的抗性,为培育小麦抗病新材料提供了基因资源。
西北农林科技大学已毕业博士舒伟学为论文第一作者,王晓杰教授和汤春蕾研究员为通讯作者,康振生教授参与并指导了该研究。研究得到了国家自然科学基金项目(32225041, 32272511)、农业生物育种国家科技重大专项(2023ZD04070)等项目资助。
1.HASP215 是 Pst重要毒性效应蛋白
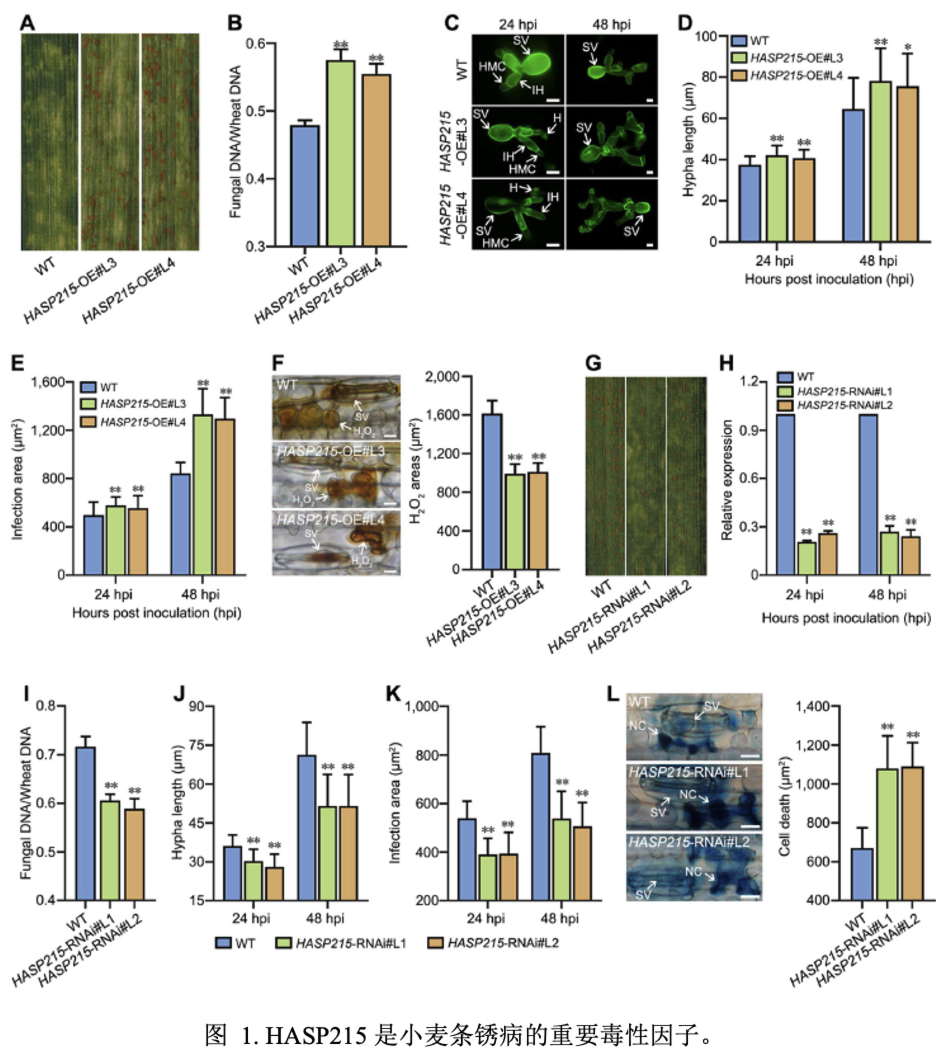
我们利用农杆菌介导的小麦遗传转化体系创制了稳定过表达HASP215的小麦转基因材料。接种CYR23后,HASP215-OE植株在产生过敏性坏死反应的同时产生少量的夏孢子堆;条锈菌侵染点周围活性氧产生面积减少,TaPR1和TaPR2表达下降,条锈菌菌丝长度与侵染面积均增加,生物量升高,表明在小麦中过表达HASP215抑制条锈菌激发的免疫反应,促进条锈菌侵染。HASP215-RNAi小麦接种CYR32,条锈菌产孢量明显减少,条锈菌菌丝长度和菌丝面积减小,生物量降低,细胞坏死面积增加,TaPR1和TaPR2表达升高,表明沉默HASP215增强了小麦对条锈菌抗性,抑制了条锈菌的致病力。结果表明,HASP215为显著影响条锈菌致病性的重要效应蛋白。
2. HASP215靶向小麦抗性正调控因子TaMKK2
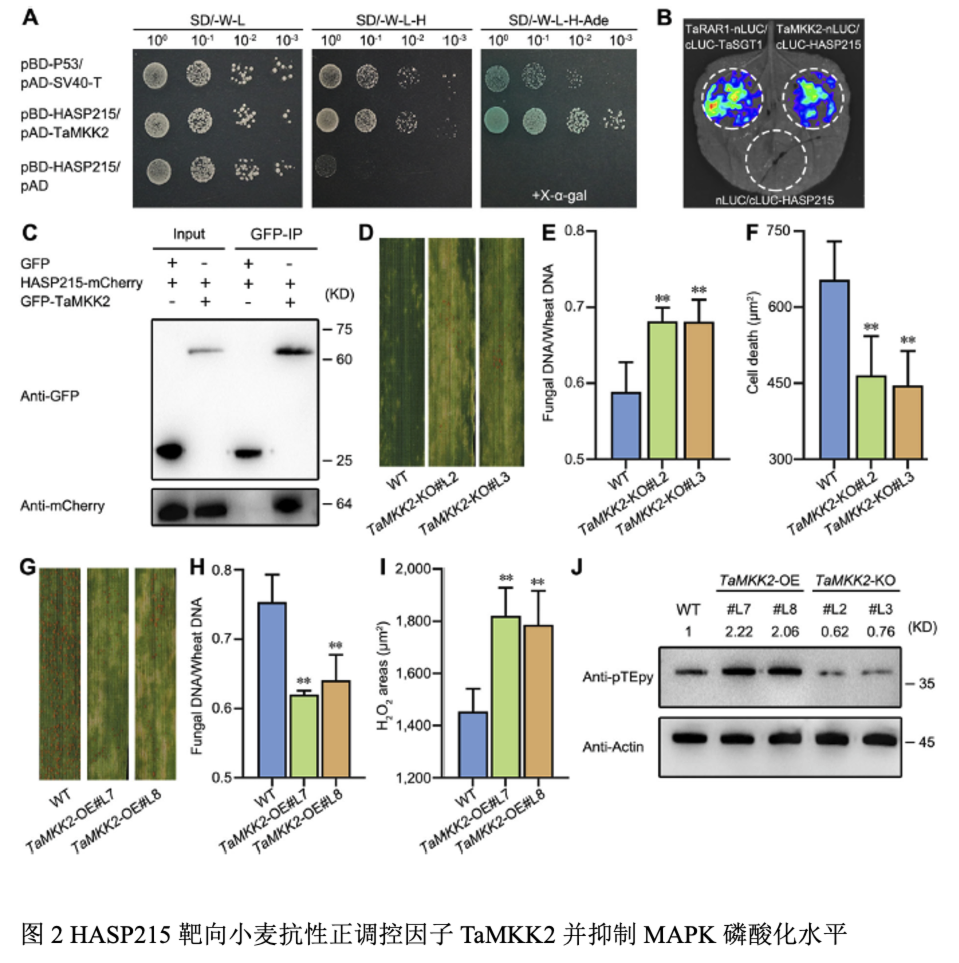
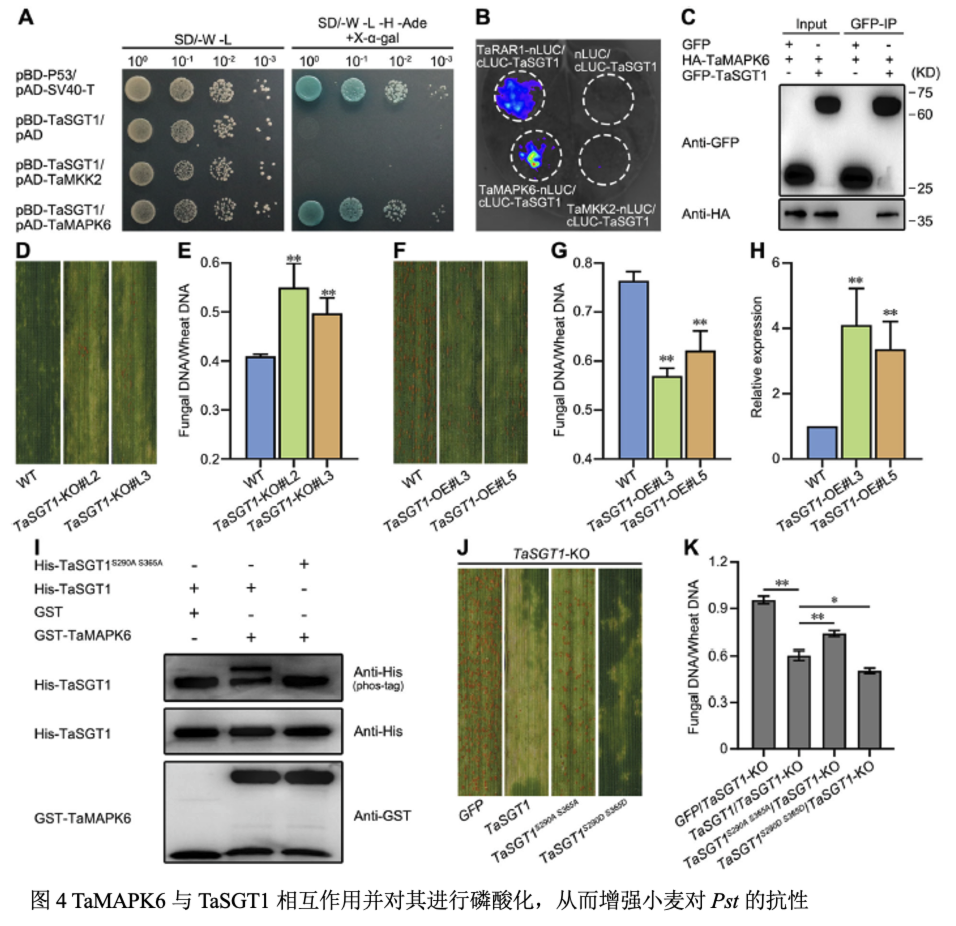
利用免疫共沉淀与质谱技术筛选HASP215的候选靶标,并利用酵母双杂交实验,荧火素酶互补实验和免疫共沉淀实验表明HASP215与小麦TaMKK2互作。利用RISPR-Cas9编辑技术创制了TaMKK2-KO突变体转基因材料,接种CYR23后,TaMKK2-KO叶片上产生少量夏孢子,菌丝长度和侵染面积增加。转基因TaMKK2-OE小麦材料接种CYR32,植株叶片条锈菌产孢量显著减少,条锈菌的生长发育受到抑制,小麦MAPK磷酸化水平增加,活性氧面积增加,TaPR1、TaPR2和TaWRKY53表达上调。在TaMKK2-OE小麦中瞬时表达HASP215则抑制MAPK磷酸化水平,降低防御相关基因表达,削弱TaMKK2-OE抗病性。结果表明TaMKK2正调控小麦对条锈菌的抗性,效应子HASP215抑制TaMKK2对条锈菌的抗性。
3. HASP215抑制TaMKK2与TaMAPK6的互作
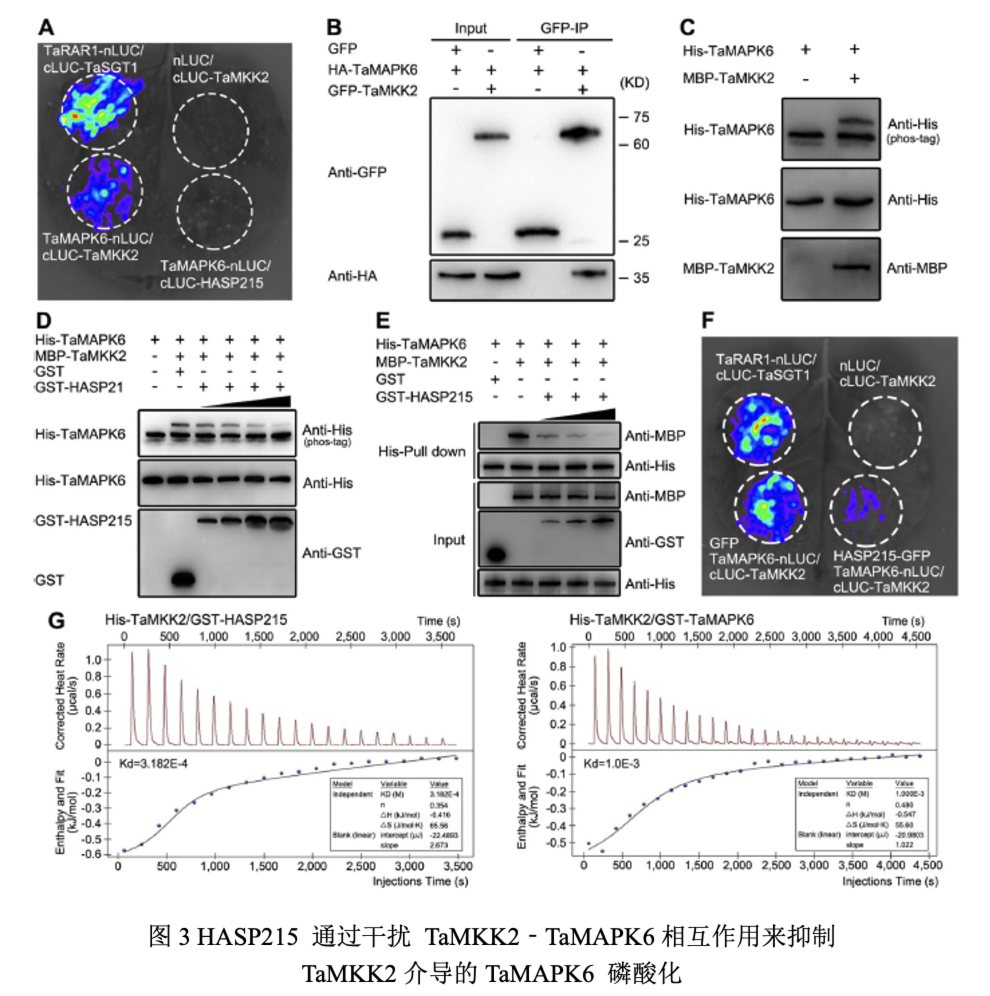
利用IP-MS筛选TaMKK2的互作靶标,并通过萤光素酶互补实验和Co-IP实验表明TaMKK2与TaMAPK4/6互作。TaMKK2能磷酸化TaMAPK4/6,而HASP215能够抑制TaMKK2对TaMAPK4/6的磷酸化,且随着HASP215蛋白量增加TaMAPK4/6磷酸化条带减弱。ITC实验表明HASP215与TaMKK2的结合亲和力高于TaMKK2与TaMAPK4/6的结合亲和力,且Pull-down实验表明随着HASP215蛋白量的增加TaMKK2与TaMAPK4/6互作减少。
4. TaMAPK6磷酸化TaSGT1促进小麦对Pst的抗性
利用酵母双杂交实验、荧火素酶互补实验及免疫共沉淀实验表明TaMAPK6能与TaSGT1互作,TaMKK2、TaMAPK4不能与TaSGT1互作。体外磷酸化实验表明TaMAPK6磷酸化TaSGT1,S290和S365是TaSGT1的关键磷酸化位点,突变后不能被TaMAPK6磷酸化。TaSGT1-OE转基因株系接种条锈菌CYR32后孢子堆数量减少,活性氧面积和细胞坏死面积显著增加,增强对条锈病抗性;TaSGT1-KO小麦接种CYR23后,产生了少量夏孢子,条锈菌的生物量与侵染面积显著增加。TaSGT1-KO转基因株系中瞬时表达TaSGT1S290AS365A(磷酸化失活)和TaSGT1S290DS365D(磷酸化激活),发现TaSGT1S290AS365A表达的TaSGT1-KO植株接种CYR23产生少量夏孢子,而TaSGT1S290DS365D瞬时表达则产生明显过敏性坏死,防卫相关基因表达升高,表明TaSGT1模拟磷酸化增强了小麦对条锈菌的抗性。
5.TaSGT1的磷酸化增强核定位促进植物免疫
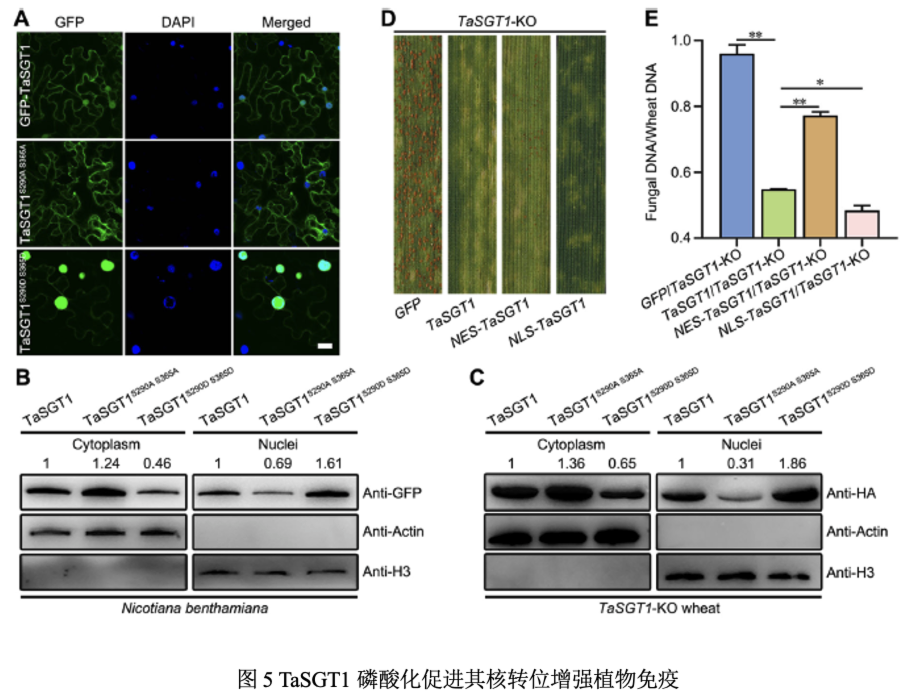
GFP-TaSGT1定位于细胞质和细胞核中,GFP-TaSGT1S290AS365A定位于细胞质和核膜,GFP-TaSGT1S290DS365D定位于细胞质和细胞核但其细胞核中的荧光信号较强。Western blot结果表明:与GFP-TaSGT1相比,GFP-TaSGT1S290AS365A在细胞质中的蛋白含量更高,GFP-TaSGT1S290DS365D在细胞核中的蛋白含量更高。TaSGT1-OE小麦接种CYR23后细胞核TaSGT1量较未接菌植株增加。在TaSGT1-OE小麦中瞬时表达HASP215则降低细胞核中TaSGT1蛋白水平,削弱小麦抗性。在TaSGT1-KO小麦中瞬时表达含有核定位信号的TaSGT1,产生过敏性坏死,瞬时表达含有核输出信号的TaSGT1的TaSGT1-KO植株,抗病性有所增强,但仍有少量夏孢子堆产生。因此TaSGT1细胞核定位增强小麦对条锈病抗性。
小麦族多组学网站:http://wheatomics.sdau.edu.cn
投稿、合作等邮箱:shengweima@icloud.com
(转自:小麦研究联盟)